Eriocalyxin B inhibits proliferation and induces apoptosis through down-regulation of Bcl-2 and activation of caspase-3 in human bladder cancer cells
Abstract
Eriocalyxin B (EriB) has been shown to possess promising anti-cancer potential against various cancer cells. However, its effect against bladder cancer cells remained unexplored. In this study, for the first time, we investigated the effects of EriB on cell proliferation, cell cycle, and apoptosis in bladder cancer T24 cells. In order to examine the effects of EriB on cell proliferation, cell cycle, and apoptosis, we performed MTT assay, flow cytometric analysis and Western blot. The results revealed that EriB decreased the cell viability of T24 cells. Flow cytometric analysis demonstrated that EriB markedly induced apoptosis of T24 cells and arrested cell cycle at G2/M phase in a dose-dependent manner. Further characterization showed that EriB involved in the down-regulation of Bcl-2 and activation of caspase-3 before culminating in apoptosis in EriB-treated T24 cells. These in vitro results suggested that EriB should be further examined for in vivo activity in human bladder cancer.
Introduction
Natural products provide many promising sources of potential anti-cancer agents and several lead structures in the past decades, which were originally isolated from plants such as paclitaxel, camptothecin, vinca alkaloids, and etoposide have potential application in cancer chemotherapy, therefore, plants are considered as one of the most vital sources for the development of novel anti-cancer drugs (Amin et al., 2009; Cragg and Newman, 2005). Diterpenoids constitute a vast class of isoprenoid natural products, which are found mainly in plants and fungi and also been found in marine organisms and insects as well (Garcia et al., 2007; Hanson, 2004). Eriocalyxin B (EriB) is a natural ent-kaurene diterpene compound isolated from Isodon eriocalyx var. laxiflora, a herb of the Labiatae family distributed in the South-West China and has been reported to have wide spectrum of biological effects, including anti-inflammatory and antibacterial agent in local folk medicine (Ikezoe et al., 2003; Wang et al., 2007). Furthermore, EriB has antiproliferative effect and induced apoptosis in cancer cells such as ovarian cancer (Leizer et al., 2010), pancreatic adenocarcinoma (Li et al., 2012) and leukemia (Wang et al., 2007; Zhang et al., 2010). However, the cytotoxic effects of EriB on bladder cancer and its mechanism were still unknown.
Bladder cancer is an increasingly common and potentially lethal malignancy (Ploeg et al., 2009). In 2012, approximately 37,510 new urinary bladder cancer cases will be diagnosed and 14,880 will die in the United States (Siegel et al., 2012). Although, many new strategies have been applied to treat bladder cancer but still further work is required in order to improve the lives of patients with this disease (Clark, 2007). In the last few decades, several natural compounds have shown a great promise in treatment of cancer and prevention of metastasis; most of the researchers are interested in and are focusing on finding a cure for cancer and several published reports have identified numerous dietary and botanical natural compounds that have chemopreventive potential (Rasul et al., 2013; Rasul et al., 2012a; Rasul et al., 2012d). Therefore, novel therapeutic agents and treatment approaches are desired to improve the clinical outcome. For this purpose, natural compounds were screened against human bladder cancer T24 cells. EriB was one of them which showed potent antiproliferative effect against the malignant bladder cancer cells. Further characterization showed that EriB effectively inhibited the proliferation of T24 cells through arresting cell cycle at G2/M phase and induction of apoptosis, which is regulated by down-regulation of Bcl-2 and activation of caspase-3.
Materials and Methods
Chemical and reagents
Eriocalyxin B (Figure 1) was purchased from the BioBioPha Co., Ltd. of Pharmaceutical and Biological Products (Kunming, China). Fetal bovine serum was purchased from Hangzhou Sijiqing Biological Engineering Materials Co., Ltd. DMEM, MTT [3'-(4,5dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide], propidium iodide (PI) and dimethyl sulfoxide (DMSO) were purchased from Sigma Chemical Company (St. Louis, USA). Annexin V-FITC Apoptosis Detection Kit and cleaved caspase-3 antibody were purchased from Beyotime Institute of Biotechnology (Shanghai, China). Rabbit polyclonal antihuman Bcl-2 antibody was purchased from Wuhan Boster Biological Technology Co., Ltd. Mouse anti-beta-actin and anti-rabbit antibodies were purchased from Santa Cruz Biotechnology. Ponceou and cell lysis buffer for Western and IP were purchased from Bio SS Beijing. Rho-123 was purchased from Eugene Co. (Oregon, U.S.A.).
Figure 1: Structure of Eriocalyxin B (EriB).
Cell culture
Human bladder cancer T24 cells were propagated in DMEM nutrients mixture supplemented with 10% FBS and antibiotics at 37°C in a humidified atmosphere with 5% CO2 and 95% air. Cells were seeded in 10 cm culture dish and allowed to grow to approximately 70% confluence before experimentation.
Cell proliferation assay
The cytotoxic effects of the EriB on the cells were determined by MTT assay as described previously (Rasul et al., 2011a; Rasul et al., 2011b). Briefly, T24 cells were seeded at a density of 1 x 104 cells per well in 96-well plates and were allowed to grow overnight. Cells were incubated with 100 µL of complete culture medium containing 0.1, 1, 3, 5, 10, and 30 µM of EriB. After incubation for 24 hours, growth of cells was determined by adding 10 µL MTT (5 mg/mL in phosphate buffered saline) to each well and incubated for 4 hours. After removal of the medium, 150 µL DMSO was added to each well and shaken carefully. The absorbance was read at a wavelength of 570 nm in a plate reader (ELX 800, BIO-TEK Instruments Inc.). The growth curve was plotted against mean values which were calculated using the following equation:
I% = [A570 (control)-A570 (treated)]/A570 (control) x 100
Flow cytometric analysis of cell cycle
For cell analysis, T24 cells were seeded in 12-well plates and then treated with 3 and 5 µM of EriB for 24 hours. After treatments, the percentages of cells in the different phases of cell cycle were evaluated by determining the DNA content after propidium iodide (PI) staining (Rasul et al., 2012b). Briefly, cells were washed with PBS, trypsinized and centrifuged at 1,000 rpm at 4°C for 5 min. Pellets were fixed overnight in 70% cold ethanol. After fixation, cells were washed twice with PBS and incubated in PBS containing RNase (1 mg/mL) for 10 min at room temperature. Finally, samples were stained with propidium iodide (1 mg/mL) for 30 min at 4°C. Data acquisition was done by flow cytometry (EPICSXL-MCL, Beckman Coulter, US) using Cell Quest software.
Flow cytometric determination of apoptosis
The apoptotic rate of T24 cells was examined by flow cytometry using annexin V-FITC/PI staining. Briefly, T24 cells were cultured in 6-well plates and allowed to attach overnight. Cells were treated with 3 and 5 uM of EriB for 24 hours. Then cells were collected, washed and resuspended in PBS. Apoptotic cell death was measured by double staining annexin V-FITC and PI using the Annexin V-FITC apoptosis detection kit (Beyotime Biotechnology Shanghai, China) according to the manufacturer's instructions. Flow cytometric analysis was performed immediately after staining. Data acquisition and analysis were performed by flow cytometry using Cell Quest software.
Western blotting
To reveal the mechanism of the apoptotic effect of EriB, Western blotting was done for apoptotic related proteins as previously described (Rasul et al., 2012c; Rasul et al., 2012d). Briefly, T24 cells were incubated with 3 and 5 µM of EriB for indicated time. Cells were trypsinized, collected in 1.5 mL centrifuge tube and washed with PBS. The cell pellets were resuspended in lysis buffer and were lysed on ice for 30 min. After centrifugation for 15 min, the supernatant fluids were collected and the protein content of the supernatant was measured by the NanoDrop 1000 spectrophotometer (Thermo scientific, USA). The protein lysates were separated by electrophoresis on 12% SDS-polyacrylamide gel and transferred to a PVDF membrane (Amersham Biosciences, Piscataway, NJ). The membranes were soaked in blocking buffer (5% skimmed milk) for 2 hours. To probe for Bcl-2, Caspase-3, and beta-actin; membranes were incubated overnight at 4°C with relevant antibodies, followed by appropriate HRP conjugated secondary antibodies and ECL detection.
Statistical analysis of data
For the statistical analysis of data, comparisons between results from different groups were analyzed with SPSS for Window Version 15.0. Student's t-test was employed to determine the statistical significance of the difference between different experimental groups and control group. p<0.05 value was defined as statistically significant. All experiments were repeated at least three times. Data were presented as mean ± standard deviation (SD).
Result and Discussion
The investigation was initiated with screening of natural compounds against T24 human bladder cancer cells. We found that EriB exhibited cytotoxic effects on the growth of T24 cells. EriB is a natural compound that belongs to a diterpene family. We determined the effects of EriB on the growth of T24 human bladder cancer cells by quantifying the viable cells using MTT assay. EriB inhibited the growth of T24 human bladder cancer cells in a dose-dependent manner (Figure 2). Morphological changes were observed under phase contrast microscopy after treating cells with 3 and 5 uM of EriB. There was a significant decrease in the number of T24 cells treated with EriB as compared to the control group. Furthermore, the cells become round-shaped and poorly adhered to the cultured plates while the control group cells showed a typical polygonal and cobblestone monolayer appearance and remained firmly attached to cultured plates (data not shown). The results indicated that EriB induced growth inhibition of T24 cells, in addition to other type of cancer cells previously reported including ovarian cancer (Leizer et al., 2010), pancreatic adenocarcinoma (Li et al., 2012) and leukemia (Wang et al., 2007; Zhang et al., 2010).
Figure 2: The effects of EriB on viability of T24 cells. T24 cells were treated with various concentrations of EriB for 24 hours. Cell death was determined by using MTT assay. Data shown are means ± SD (n = 3)
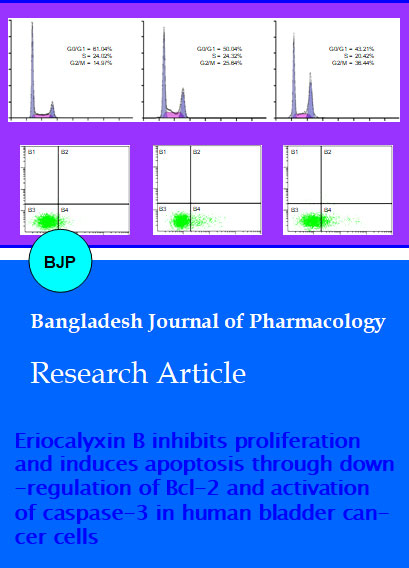
There are several mechanisms which control the cell cycle to ensure the correct cell division. It is well known that progression of cell cycle is maintained by different check points in normal cells and the transition from one cell cycle phase to another occurs in an orderly fashion. In cancerous cells, some basic modifications occurred in the genetic control of cell division, resulting in an uncontrolled cell proliferation. As the deregulation of cell cycle progression is the hallmark of cancer; thereby cell cycle regulation could be a potential and effective strategy for the treatment of cancer (Grana and Reddy, 1995; Vermeulen et al., 2003). Therefore, we analyzed effect of EriB on cell cycle progression of T24 cells. It was found that EriB arrested cell cycle at G2/M phase, which supported the previous reported results in pancreatic adenocarcinoma (Li et al., 2012). The percentage of accumulation of cells in the G2/M phase was increased from 15.97% in control group to 25.64%, and 38.48% in the cells treated with 3 and 5 uM of EriB respectively for 24 hours (Figure 3). These findings revealed that G2/M phase cell cycle arrest was one of the mechanisms through which EriB induces cytotoxicity in T24 cells. A number of recent studies have shown that by arresting the cell division at certain checkpoints in the cell cycle, several chemotherapeutic and chemopreventive agents have demonstrated potential antiproliferative effects (Rasul et al., 2013; Rasul et al., 2012b).
Figure 3: Effect of EriB on the cell cycle distribution. T24 cells were treated with 0, 3 and 5 uM of EriB for 24 hours and then they were stained with PI for flow cytometric analysis. Histograms show number of cells/channel (y-axis) vs. DNA content (x-axis) and the values indicate the percentage of cells in the indicated phases of cell cycle. The data shown are representative of three independent experiments with the similar results. ap<0.05 and bp<0.01 compared with the control
There are various modes of cell death such as apoptosis, autophagy and necrosis (Leist and Jaattela, 2001). Apoptosis is most organized, well fashioned, and systematic mode of cell death, in which cells themselves play an active role in their own death (Elmore, 2007; Hengartner, 2000). The normal cellular signals for regulation of their growth are lost in the cancerous cells due to various mutations, preventing the cells from apoptosis and cell growth to uncontrolled status (Hanahan and Weinberg, 2000). The regulation of apoptosis is, therefore, most important in the treatment of cancer (Fulda, 2010; Lawen, 2003; Reed, 2002). The chemopreventive agents, which can treat the cancer effectively, have potential to restore the natural signaling apoptotic pathway (Reed, 1999). It is well known that various chemopreventive agents cause cell death through induction of apoptosis in different cancer cells (Srivastava and Gupta, 2006; Xu et al., 2009). We studied whether EriB inhibits cell growth in T24 cells through the induction of apoptosis. EriB-induced apoptosis was determined by flow cytometric analysis.
For flow cytometric analysis, cells were seeded in the 12 well plates. After incubation of cells without (control) or with EriB for 24 hours, cells were collected in centrifuged tubes and stained with annexin V-FITC and PI double staining as described in material and methods part. The results of flow cytometric analysis showed that rates of apoptosis were 17.6 ± 2.1 and 24.9 ± 2.5% in the cells treated with 3 and 5 uM of EriB respectively for 24 hours as compared to 5.9 ± 1.0% in the control cells (Figure 4). EriB-induced apoptosis in T24 cells was consistent with previously reported studies in ovarian cancer (Leizer et al., 2010), pancreatic adenocarcinoma (Li et al., 2012) and leukemia (Wang et al., 2007; Zhang et al., 2010).
Figure 4: Effects of EriB on apoptosis in T24 cells. (A) T24 cells were treated with 0, 3 and 5 uM of EriB for 24 hours. Then they were stained with Annexin V-FITC/PI and were analyzed in flow cytometry. Cell populations shown in the lower right (Annexin V+/PI-) represents early apoptotic cells, upper right (Annexin V+/PI+) represents late apoptotic cells. (B) The values indicate the percentage of apoptotic cells treated without (control) and with (3 and 5 uM) EriB for 24 hours. The data shown are representative of three independent experiments with the similar results. ap<0.05 and bp<0.01 compared with the control
Mitochondria play a fundamental role in the regulation of apoptotic cell death and consist of various pro-apoptotic proteins and cytochrome C. As highlighted earlier, apoptosis involves a dysfunction of mitochondrial membrane integrity, which leads to cell death (Jeong and Seol, 2008). Previously it has been documented that disintegration of the mitochondrial membrane potential and the redistribution of cytochrome C are crucial actions in the apoptotic cascade (Kluck et al., 1997; Wang, 2001). Cytochrome C plays central role in mitochondrial mediated apoptosis. Upon the attenuation of mitochondrial transmembrane potential, Cytochrome C releases from the mitochondria into the cytosol (Kluck et al., 1997). Once released into the cytosol, cytochrome C binds to form an "apoptosome" of Apaf-1, cytochrome C, and caspase-9, which subsequently cleaves the effecter caspase-3 (Ricci and Zong, 2006).
The caspases are a family of proteins related to cysteine proteases that is one of the focal executors of the apoptotic process via triggering the death receptors and mitochondrial pathways to accomplish the programmed cell death (Cohen, 1997). Caspases are present in the form of inactive zymogens those are activated during apoptosis. Among them, caspase-3 is a frequently activated death protease, catalyzing the specific cleavage of many key cellular proteins (Adams, 2003; Porter and Janicke, 1999). In order to determine effects of EriB on expression of caspase-3, Western blotting was done. The results showed that procaspase-3 was cleaved to its active fragments in treated cells with 3 and 5 µM of EriB after 24 hours as compared to that of control cells (Figure 5). These findings are supported by previous studies (Kanno et al., 2008; Lee et al., 2001; Park et al., 2001). These results markedly showed that EriB induced caspase-dependent cell death in T24 cells.
Figure 5: The effects of EriB on expression levels of apoptosis related proteins. Expression levels of Bcl-2 and caspase-3 in T24 cells treated without (control) and with (3 and 5 uM) EriB for 24 hours were monitored by Western blot assay. beta-actin was used as loading control. Western blots are representative of three independent experiments
The release of cytochrome c from the mitochondria into cytosol is coupled with the ratio of Bax/Bcl-2 proteins (Kluck et al., 1997). Bcl-2 family proteins are mostly involved in the mitochondrial apoptotic pathway (Reed, 1998). Furthermore, Bcl-2 family plays a central role in activation of Caspases (Burlacu, 2003). Bcl-2 and Bax work in an antagonistic manner, Bcl-2 is anti-apoptotic while Bax is proapoptotic member of Bcl-2 family and Bcl-2 forms heterodimers with Bax. Bax inserts into outer membrane of mitochondria under stress conditions, as a results transmembrane permeability become higher which facilitate the release of cytochrome C due to formation of pores on the mitochondrial outer membrane (Adams and Cory, 2007; Danial, 2007). The balance between these two groups is critical in the cell decision to undergo apoptosis or not (Mallat and Tedgui, 2000). We performed Western blotting to examine the effect of EriB on the expression of Bcl-2. EriB was observed to be involved in the down regulation of Bcl-2 in dose-dependent manner (Figure 5). These results are similar with previously reported studies in pancreatic adenocarcinoma (Li et al., 2012).
Conclusion
To conclude, EriB induced apoptosis of T24 human bladder cancer cells accompanied by a G2/M phase cell cycle arrest. Further characterization showed that EriB involved in the down-regulation of Bcl-2 and activation of caspase-3 before culminating in apoptosis in EriB-treated T24 cells. These in vitro results suggested that EriB should be further examined for in vivo activity and molecular mechanism in human bladder cancer.
References
Adams JM. Ways of dying: Multiple pathways to apoptosis. Genes Dev. 2003; 17: 2481-95.
Adams JM, Cory S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene 2007; 26: 1324-37.
Amin ARMR, Kucuk O, Khuri FR, Shin DM. Perspectives for cancer prevention with natural compounds. J Clin Oncol. 2009; 27: 2712-25.
Burlacu A. Regulation of apoptosis by Bcl-2 family proteins. J Cell Mol Med. 2003; 7: 249-57.
Clark PE. Bladder cancer. Curr Opin Oncol. 2007; 19: 241-47.
Cohen GM. Caspases: The executioners of apoptosis. Biochem J. 1997; 326: 1-16.
Cragg GM, Newman DJ. Plants as a source of anti-cancer agents. J Ethnopharmacol. 2005; 100: 72-79.
Danial NN. BCL-2 family proteins: Critical checkpoints of apoptotic cell death. Clin Cancer Res. 2007; 13: 7254-63.
Elmore S. Apoptosis: A review of programmed cell death. Toxicol Pathol. 2007; 35: 495-516.
Fulda S. Evasion of apoptosis as a cellular stress response in cancer. Int J Cell Biol. 2010; 2010: 1-6.
Garcia PA, de Oliveira AB, Batista R. Occurrence, biological activities and synthesis of kaurane diterpenes and their glycosides. Molecules 2007; 12: 455-83.
Grana X, Reddy EP. Cell cycle control in mammalian cells: Role of cyclins, cyclin dependent kinases (CDKs), growth suppressor genes and cyclin-dependent kinase inhibitors (CKIs). Oncogene 1995; 11: 211-19.
Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000; 100: 57-70.
Hanson JR. Diterpenoids. Nat Prod Rep. 2004; 21: 785-93.
Hengartner MO. The biochemistry of apoptosis. Nature 2000; 407: 770-76.
Ikezoe T, Chen SS, Tong XJ, Heber D, Taguchi H, Koeffler HP. Oridonin induces growth inhibition and apoptosis of a variety of human cancer cells. Int J Oncol. 2003; 23: 1187-93.
Jeong SY, Seol DW. The role of mitochondria in apoptosis. BMB Rep. 2008; 41: 11-22.
Kanno S, Kitajima Y, Kakuta M, Osanai Y, Kurauchi K, Ujibe M, Ishikawa M. Costunolide-induced apoptosis is caused by receptor-mediated pathway and inhibition of telomerase activity in NALM-6 cells. Biol Pharm Bull. 2008; 31: 1024-28.
Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The release of cytochrome c from mitochondria: A primary site for Bcl-2 regulation of apoptosis. Science 1997; 275: 1132-36.
Lawen A. Apoptosis-an introduction. Bioessays 2003; 25: 888-96.
Lee MG, Lee KT, Chi SG, Park JH. Costunolide induces apoptosis by ROS-mediated mitochondrial permeability transition and cytochrome C release. Biol Pharm Bull. 2001; 24: 303-06.
Leist M, Jaattela M. Four deaths and a funeral: From caspases to alternative mechanisms. Nat Rev Mol Cell Biol. 2001; 2: 589-98.
Leizer AL, Alvero AB, Fu HH, Holmberg JC, Cheng YC, Silasi DA, Rutherford T, Mor G. Regulation of inflammation by the NF-kappaB pathway in ovarian cancer stem cells. Am J Reprod Immunol. 2010; 65: 438-47.
Li L, Yue GG, Lau CB, Sun H, Fung KP, Leung PC, Han Q, Leung PS. Eriocalyxin B induces apoptosis and cell cycle arrest in pancreatic adenocarcinoma cells through caspase- and p53-dependent pathways. Toxicol Appl Pharmacol. 2012; 262: 80-90.
Mallat Z, Tedgui A. Apoptosis in the vasculature: Mechanisms and functional importance. Br J Pharmacol. 2000; 130: 947-62.
Park HJ, Kwon SH, Han YN, Choi JW, Miyamoto K, Lee SH, Lee KT. Apoptosis-Inducing costunolide and a novel acyclic monoterpene from the stem bark of Magnolia sieboldii. Arch Pharm Res. 2001; 24: 342-48.
Ploeg M, Aben KKH, Kiemeney LA. The present and future burden of urinary bladder cancer in the world. World J Urol. 2009; 27: 289-93.
Porter AG, Janicke RU. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999; 6: 99-104.
Rasul A, Bao R, Malhi M, Zhao B, Tsuji I, Li J, Li X. Induction of apoptosis by costunolide in bladder cancer cells is mediated through ROS generation and mitochondrial dysfunction. Molecules 2013; 18: 1418-33.
Rasul A, Ding C, Li X, Khan M, Yi F, Ali M, Ma T. Dracorhodin perchlorate inhibits PI3K/Akt and NF-kappaB activation, up-regulates the expression of p53, and enhances apoptosis. Apoptosis 2012a; 17: 1104-19.
Rasul A, Khan M, Yu B, Ma T, Yang H. Xanthoxyletin, a coumarin induces S phase arrest and apoptosis in human gastric adenocarcinoma SGC-7901 cells. Asian Pac J Cancer Prev. 2011a; 12: 1219-23.
Rasul A, Song R, Wei W, Nishino Y, Tsuji I, Li X, Li J. Tubeimoside-1 inhibits growth via the induction of cell cycle arrest and apoptosis in human melanoma A375 cells. Bangladesh J Pharmacol. 2012b; 7: 150-56.
Rasul A, Yu B, Khan M, Zhang K, Iqbal F, Ma T, Yang H. Magnolol, a natural compound, induces apoptosis of SGC-7901 human gastric adenocarcinoma cells via the mitochondrial and PI3K/Akt signaling pathways. Int J Oncol. 2012c; 40: 1153-61.
Rasul A, Yu B, Yang LF, Ali M, Khan M, Ma T, Yang H. Induction of mitochondria-mediated apoptosis in human gastric adenocarcinoma SGC-7901 cells by kuraridin and Nor-kurarinone isolated from Sophora flavescens. Asian Pac J Cancer Prev. 2011b; 12: 2499-504.
Rasul A, Yu B, Zhong L, Khan M, Yang H, Ma T. Cytotoxic effect of evodiamine in SGC-7901 human gastric adenocarcinoma cells via simultaneous induction of apoptosis and autophagy. Oncol Rep. 2012d; 27: 1481-87.
Reed JC. Bcl-2 family proteins. Oncogene 1998; 17: 3225-36.
Reed JC. Dysregulation of apoptosis in cancer. J Clin Oncol. 1999; 17: 2941-53.
Reed JC. Apoptosis-based therapies. Nat Rev Drug Discov. 2002; 1: 111-21.
Ricci MS, Zong WX. Chemotherapeutic approaches for targeting cell death pathways. Oncologist 2006; 11: 342-57.
Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer J Clin. 2012; 62: 10-29.
Srivastava JK, Gupta S. Tocotrienol-rich fraction of palm oil induces cell cycle arrest and apoptosis selectively in human prostate cancer cells. Biochem Biophys Res Commun. 2006; 346: 447-53.
Vermeulen K, Van Bockstaele DR, Berneman ZN. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2003; 36: 131-49.
Wang L, Zhao WL, Yan JS, Liu P, Sun HP, Zhou GB, Weng ZY, Wu WL, Weng XQ, Sun XJ, Chen Z, Sun HD, Chen SJ. Eriocalyxin B induces apoptosis of t(8;21) leukemia cells through NF-kappaB and MAPK signaling pathways and triggers degradation of AML1-ETO oncoprotein in a caspase-3-dependent manner. Cell Death Differ. 2007; 14: 306-17.
Wang X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001; 15: 2922-33.
Xu Y, Chiu JF, He QY, Chen F. Tubeimoside-1 exerts cytotoxicity in HeLa cells through mitochondrial dysfunction and endoplasmic reticulum stress pathways. J Proteome Res. 2009; 8: 1585-93.
Zhang YW, Jiang XX, Chen QS, Shi WY, Wang L, Sun HD, Shen ZX, Chen Z, Chen SJ, Zhao WL. Eriocalyxin B induces apoptosis in lymphoma cells through multiple cellular signaling pathways. Exp Hematol. 2010; 38: 191-201.