Periostin as a promising target of therapeutical intervention for colorectal cancer
Abstract
The expression of periostin in the tissue of colorectal cancer patients and its correlation with clinical features were studied. Periostin expression was ~4 fold up-regulated in cancer tissues compared to adjacent non-cancerous tissues. Serum levels of periostin in patients were significantly elevated to 32.6 ± 10.8 ng/mL vs 20.4 ± 11.1 ng/mL in healthy volunteers. Higher preoperative serum periostin levels in patients were closely related to advanced-stage disease (stage III/IV), distant and lymph nodes metastasis. High level of periostin expression was detected in the SW480 human colon carcinoma cells, and could be down-regulated by small interfering RNAs (siRNA). The siRNA-mediated knockdown of periostin arrested the cell cycle at G0/G1 phase and induced apoptotic cell death in the SW480 cells. In conclusion, periostin is a promising prognostic and therapeutic target for colorectal cancer.
Introduction
Colorectal cancer is the third most common malignancy in developed countries, which accounts for the second most frequent cause of cancer mortality worldwide (Herszenyi et al., 2010; American Cancer Society. 2013-2014; Hwang, 2013). There are 1-2 million new cases and bring about 600,000 mortalities each year (Brenner et al., 2013; Rose et al., 2014).
Despite recent progresses in diagnosis of colorectal cancer and in its management (Dattatreya, 2013; Price et al., 2013; Khoshbaten et al., 2014), the number of mortalities is still progressively increasing every year. Many patients with localized colorectal cancer have micrometastatic disease are usually miss diagnosed, which results in disease recurrence and mortality several years after diagnosis (Noura et al., 2002). So, there is an urgent need for identifying novel serum biomarkers related to tumor aggressiveness and metastasis (Lech et al., 2014).
Periostin was originally identified in osteoblast cells as an important regulator of bone and tooth formation and maintenance (Takeshita et al., 1993; Horiuchi et al., 1999). It is now determined that periostin is a heterofunctional secreted extracellular matrix protein that is associated with cellular adhesion and movement during embryonic development (Conway et al., 2008; Sorocos et al., 2011).
Periostin is also highly expressed at sites of injury (Conway et al., 2014), or inflammation within the adult organism where it plays critical roles in embryonic development (Brenner et al., 2013; Rose et al., 2014). For example, periostin is predominantly expressed in endothelial-mesenchymal transformation valve tissues and related supporting apparatus in both embryonic and adult where it regulates post-endothelial-mesenchymal transformation atrioventricular valve differentiation (Norris et al., 2008). Periostin is also frequently over-expressed in various types of human cancer tissues (Luo et al., 2013) and promotes invasion, angiogenesis and metastasis of these tumors. Ben and colleagues (Ben et al., 2009) analyzed the periostin levels of serum and tissue samples of colorectal cancer patients by ELISA and immunohistochemistry methods, and found up-regulation of periostin expression in most sample. A recent report indicated that periostin was related to the liver metastasis of colorectal cancer (Wu et al., 2013).
To further understand the roles of periostin in colorectal cancer progression, we evaluated periostin expression in colorectal cancer tissues and adjacent non-cancerous tissues by proteomics method. We also determined serum periostin levels of both colorectal cancer patients and healthy volunteers by ELISA. We then determined the expression of periostin in SW480 cell lines by immunofluorescence staining assay. Next, the SW480 cells with periostin expression were used for transfecting siRNA-periostin lentiviral particles. The efficacy of transfecting siRNA lentiviral particles was evaluated by immunofluorescence, Western blotting and real-time PCR. The effect of silencing periostin expression by siRNA on proliferation of sw480 cells was determined by MTT assay. Cell cycle distributions and apoptosis profiles were evaluated by flow cytometry assay.
Materials and Methods
Tissue specimens
Cancer tissues and adjacent non-cancerous tissues (with 10 cm distance from tumor edge) were obtained from 65 colorectal cancer patients, at the first hospital of Jiamus University between February 2010 and February 2012. All patients were diagnosed with Dukes C phase colorectal cancer and received no therapy prior to surgery. The tissues were frozen and stored at -80°C until use.
Protein extraction
The frozen tissues were cut into small pieces and homogenized in liquid nitrogen. The homogenized samples were then extracted with ice-cold protein extraction kit (Readyprep protein extraction kit, BioRad, USA) for 20 min. Supernatant was collected by centrifugation at 12000 x g for 4 min. The protein concentrations were determined using a BCA protein Assay Kit.(Rockford, IL, USA).The supernatants were stored at -80°C before use.
Proteolysis
The supernatants were thawed at room temperature and dissolved in 3-5 volumes (v/v) NH4HCO3 (50 mM) solution. For protein solubilization and denaturation, the samples were incubated with 20 mM DTT-containing NH4HCO3 at 56°C for 1 hour, and then with 50 mM IAM-containing NH4HCO3 at room temperature for 30 min, sequentially. The samples were then incubated with gold grade trypsin (Promega, Madison, WI) overnight at 37°C.
LC-MS/MS analysis
The samples were dissolved in 0.1% formaldehyde. The samples (50 ug) were loaded on a C18 trap column (C18 PepMap, 300 um ID x 5 mm, 5 um particle size,100 Angstrom pore size; Dionex, Amsterdam, The Netherlands) and eluted onto a SCX capillary column (100 mm x 320 um id). The ammonia acetate concentration of elution steps for SCX capillary column were 0 mM, 25 mM, 75 mM, 100 mM and 1M. The eluted peptides were then transferred onto a RP capillary column (PepMap C18, 75 um ID Angstrom 150 mm, 3 um particle and 100 Angstrom pore size; Dionex, Amsterdam, The Netherlands). The elution gradient for RP column was from 5% to 30% buffer B (0.1% formic acid, 99.9% acetonitrile) over 3 hours at a flow rate of 2 uL/min. MS data were acquired in a survey scan from 400-2000 amu (1 μscans) followed by ten data-dependent MS/MS scans (10 μscans each, isolation width 3 amu, 35% normalized collision energy, dynamic exclusion for 1.5 min) on an LTQ XL electrospray ion trap mass spectrometer (ThermoFisher, San Jose, CA).
Serum periostin level was determined using Human Peristin ELISA kit (RD, USA) as manufacturer's protocol. Briefly, 96-well plates were coated with anti-periostin monoclonal antibody at 4°C, overnight. The plates were then coated with bovine serum albumin for 2 hours, at room temperature. In each well of the microplate 50 uL diluted serum sample was added and maintained at 37°C 1 hour. After washing the unbounded periostin carefully with PBS, 50 uL horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin (1:500 dilution in PBS) was added in each well and the plate was incubated at 37°C for 30 min. The absorbance (at 450 nm) of each well was determined using a microplate reader (ELX 800, BIO-TEK Instruments).
Cell culture and siRNA lentiviral particles transfection
SW480 cells were cultured in RMPI-1640 medium supplemented with 10% fetal bovine serum (FBS) and 100 U/mL of penicillin-streptomycin (Invitrogen) in a CO2 incubator at 37°C with 5% CO2 in humidified atmosphere. siRNA-PERIOSTINs were designed to target the periostin sequences (GenBank accession no. NM_001135934.1) at 1013 and 2878, respectively. The siRNA-1 sequences are listed below:
siRNA-1 forward: ccggcacgGTGACAGTATAACAGTAA ACTCGAGTTTACTGTTATACTGTCACCGTTTTTg siRNA-1 reverse: aattcaaaaacacgGTGACAGTATAACA GTAAACTCGAGATAACATACATACAAGGCTCG
siRNA-2 forward: ccggcacgAGCCTTGTATGTATGTTA TCTCGAGTTTACTGTTATACTGTCACCGTTTTTg
siRNA-2 reverse: aattcaaaaacacgAGCCTTGTATGTAT GTTATCTCGAGATAACATACATACAAGGCTCG
siRNA-1 and siRNA-2 were cloned into the GFP-labeled lentiviral expression vector GV248 to get psiRNA-1 and psiRNA-2. The recombinant vectors were transfected into SW480 cells using the gene transfection kit GenEscortTM as the manufacturer's protocol.
Quantitative polymerase chain reaction
Total RNA was isolated using trizol reagent (Invitrogen, USA) and reversely transcribed into cDNA using all-in-one first-strand cDNA synthesis kit (Genecopoeia, China) as the manufacturer's protocols. The periostin gene was quantified by real time PCR method using the real time PCR master mix kit (Genecopoeia, China), beta-actin was used as an internal marker. The primers used were as follows:
Actin-forward: 5'-CATGTACGTTGCTATCCAGGC-3'
Actin-reverse: 5'-CTCCTTAATGTCACGCACGAT-3'
Periostin-forward: 5'-CCCCGTGACTGTCTATAAGC-3'
Periostin-reverse: 5'-AAATGACCATCACCACCTTCA-3'
Immunofluorescence staining
SW480 cells were grown on glass cover slips till 80% confluent. After washing 3 times with Hank's solution, the cells were fixed with 4% paraformaldehyde (PFA) for 30 min. The cells were washed twice with PBS, incubated with 0.1%Triton X-100 for 10 min, and washed 3 times with PBS again. The cells were then blocked with 5% goat serum albumin for 1 hour, incubated with rabbit polyclonal anti-periostin antibody (SAB, 1:100) for 1 hour, followed by incubation with Alexa555-labeled goat anti-rabbit secondary antibodies (Molecular Probe, 1:200) for 1 hour at room temperature. The expression of periostin was visualized under a fluorescence microscope.
Western blotting
SW480 cells were incubated with siRNAs for 24 or 48 hours, collected, washed 3 times with PBS, and then lysed with RIPA lysis buffer (Beyotime, Shanghai, China). Supernatants of the cell lysates were collected by centrifugation at 12,000 rpm for 15 min. Proteins were fractionated using 12% SDS-PAGE and transferred to PVDF membranes according to standard protocols. The membranes were blocked with 5% non-fat milk, incubated overnight at 4°C with the primary antibody, washed 3 times with PBS, and then incubated with secondary antibody at room temperature. The intensities of target proteins were detected using enhanced chemiluminescence kit on X-ray film (Millipore Corporation, USA).
Determination of cell viability
Cell viability was determined by MTT assay as described previously (Khan et al., 2013). Briefly, SW480 cells were incubated with siRNAs in 96-well tissue culture plate for 24 or 48 hours. After the treatment, the cells were incubated with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma, USA) at 37°C for 4 hours. Following incubation, 150 μL dimethylsulfoxide (DMSO, Sigma, USA) was added to dissolve the farmazan crystals. The absorbance (OD570) of the suspension was measured by using a microplate reader (ELX 800, BIO-TEK Instruments). Percentage cell viability was calculated as follows:
Cell viability (%) = (A570 sample - A570 blank) / (A570 control - A570 blank) x 100
Flow cytometric analysis of apoptosis
SW480 cells were seeded in 6-well plates at a density of 106. After adherence, the cells were exposed to siRNAs for 48 hours. The cells were then harvested, washed 3 times with PBS, and labeled with annexin V-FITC (Beyotime, China) for 15 min in the dark. The cells were then visualized for the fluorescence of annexin V under a fluorescent microscope (Olympus 1x 71). At the end, 100 cells were counted microscopically for the percentage of live and apoptotic cells.
Flow cytometric analysis of cell cycle
SW480 cells were incubated with the siRNAs in 6-well plates for 48 hours and cell cycle profile was analyzed as described previously (Khan et al., 2012). Briefly, the cells were collected and washed with PBS 3 times. Cell pellets were fixed with ethanol (70%) at -20degree Centegrade overnight, centrifuged, washed with PBS and stained with a solution containing 50 ug/mL PI and 100 ug/mL RNase A for 30 min in the dark at room temperature. Cell cycle distribution was assayed on a flow cytometry and analyzed by Cell Quest software.
Statistical analysis
Data were expressed as mean ± SEM, and analyzed by one-way ANOVA with SPSS software, version 13.0 (SPSS Inc., USA). p values less than 0.05 were considered statistically significant.
Results
To identify candidate biomarkers for colorectal cancer, we compared the protein expression profiles of colorectal cancer tissues and adjacent normal tissues from 65 patients by LC-MS/MS method. The results indicated that 15 proteins were up-regulated in colorectal cancer as compared to the normal control, among which periostin expression was ~4 fold up-regulated in colorectal cancer tissues as compared to the matched adjacent normal tissue. Mass spectrometry results of periostin isoforms was shown in Table I.
Table I: Identification of periostin isoforms by mass spectrometry analysis
| Scan(s) | Peptide | MH+ | DeltaM | z | XC | DeltaCn |
|---|---|---|---|---|---|---|
| 10671 | R.IMGDKVASEALMK.Y | 1393.7 | 0.88 | 2 | 3.79 | 0.52 |
| 12139 | K.TTVLYECCPGYMR.M | 1650.9 | 0.69 | 2 | 2.92 | 0.50 |
| 20847 | K.GCPAVLPIDHVYGTLGIVGATTTQR.Y | 2597.9 | 1.15 | 3 | 3.00 | 0.26 |
| 21679 | K.VGLNELYNGQILETIGGK.Q | 1919.2 | 0.83 | 2 | 4.26 | 0.51 |
| 31032 | R.AAAITSDILEALGRDGHFTLFAPTNEAFEKLPR.G | 3574.0 | 0.39 | 3 | 5.83 | 0.67 |
| 31550 | K.DIVTNNGVIHLIDQVLIPDSAK.Q | 2375.7 | 0.03 | 2 | 4.52 | 0.52 |
| 33899 | R.VLTQIGTSIQDFIEAEDDLSSFR.A | 2585.8 | -0.4 | 2 | 5.14 | 0.65 |
| 33874 | K.SKESDIMTTNGVIHVVDKLLYPADTPVGNDQLLEILNK.L | 4182.7 | 0.89 | 3 | 3.62 | 0.65 |
Next, we examined the serum periostin levels of the 65 colorectal cancer patients. We found that the serum levels of periostin in colorectal cancer patients were significantly elevated to 32.6 ± 10.8 ng/mL vs 20.4 ± 11.1 ng/mL in healthy volunteers. Further analysis indicated that higher preoperative serum periostin levels in colorectal cancer patients were closely related to advanced-stage disease (stage III/IV), distant and lymph nodes metastasis, while there was no significant correlation between periostin expression and such clinical features as age or sex of patients and differentiation of tumor (Table II).
Table II: Correlation between plasma periostin concentration and clinic-pathological features in colorectal cancer
| Sample size | Periostin (ng/mL) | p | ||
|---|---|---|---|---|
| Sex | Male | 41 | 32.7 ± 11.3 | >0.05 |
| Female | 24 | 32.4 ± 10.1 | >0.05 | |
| Age | <55 | 13 | 35.1 ± 10.1 | >0.05 |
| >55 | 52 | 32.0 ± 11.8 | >0.05 | |
| Differentiated | Well/Mid-well | 48 | 31.2 ± 11.6 | >0.05 |
| Poor | 17 | 36.7 ± 9.5 | >0.05 | |
| TNM stage | I+II | 36 | 27.0 ± 10.9 | <0.01 |
| III+IV | 29 | 36.4 ± 9.3 | <0.01 | |
| Lymph nodes metastasis | N0 | 29 | 28.0 ± 10.7 | |
| N1 | 27 | 36.1 ± 9.2 | <0.01 | |
| N2 | 9 | 37.2 ± 9.5 | <0.05 | |
| Distal metastasis | M0 | 50 | 31.1 ± 10.8 | <0.05 |
| M1 | 15 | 37.8 ± 9.1 | <0.05 |
First, we determined the expression of periostin in SW480 colon carcinoma cells by using immunofluorescence staining method. We found robust expression of periostin protein in SW480 cells (Figure 1A). The periostin-expressing SW480 cells were used for transfection of siRNA-periostin lentiviral particles. The cells were transfected with two kinds of siRNAs (psiRNA-1 and psiRNA-2), and each for 24 and 48 hours, respectively. The efficiencies of the transfection were evaluated by immunofluorescence, Western blotting and real-time PCR assays. Cellular imaging provided an estimated transfection efficiency of 50-70% (Figure 1B). Next, we determined periostin mRNA and protein levels in the siRNAs transfected SW480 cells. Real-time PCR results indicated that periostin levels in SW480 cells were decreased by more than 70% after transfecting with the siRNA lentiviral particles (Figure 1C). As shown in Figure 1D, the periostin protein level was also significantly reduced as was expected. These results indicated that periostin siRNA could effectively down-regulated the expression of periostin in SW480 cells.
Figure 1: The efficiency of periostin siRNA lentiviral particles transfection in SW480 cells. A, Periostin expression in SW480 cells was determined by immunofluorescence staining method. Nuclei were stained with Hoechest33342 (blue) and periostin were stained with Alexa555-labled antibodies (red); B, Relative fluorescence intensities of siRNA levels in transfected SW480 cells, data were expressed as mean ± SEM, n=6; C, Decreased expression of periostin mRNA in siRNA-trasfected SW480 cells, data were expressed as mean ± SEM, n=6, ap<0.01; D, Decreased expression of periostin protein was confirmed by western blotting method Expression of periostin in SW480 cells were determined using Western blotting assay, beta-actin was used as loading control.
Inhibition of periostin expression induced apoptotic cell death in SW480 cells
Previous studies showed that periostin was involved in apoptosis regulation in several cancer cell lines (Liu et al., 2010; Zhu et al., 2011; Luo et al., 2013; Li et al., 2013). We (avoid) were, therefore, curious to know whether the targeted down-regulation of periostin cells could induce apoptosis in SW480 cells. We performed MTT assay to evaluate the effect of siRNAs on cell viability in SW480 cells. As shown in Figure 2, both periostin siRNAs significantly reduced cell viability in SW480 cells after 48 hours treatment, as compared with the control cells.
Figure 2: Silencing periostin inhibits proliferation in SW480 cells. The SW480 cells were transfected with p-siRNA-1, p-siRNA-2 and control GFP lentiviral particles for 24 or 48 hours, and cells viability was determined by MTT assay. Averaged cell viability values were expressed as mean ± SEM, of three independent experiments, ap<0.01 compared with the control
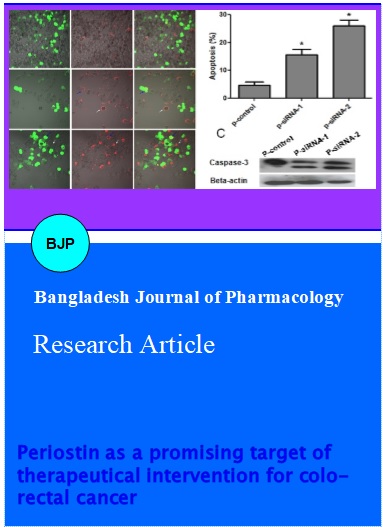
Next, we used Annexin V-FITC staining method to determine the apoptotic effects of the siRNAs in SW480 cells. After treating with p-siRNA-1 and p-siRNA-2 for 48 hours, the SW480 cells were harvested and stained with annexin V-FITC. The stained cells were then observed under fluorescence microscope for the detection of apoptosis. The data demonstrated that both p-siRNA-1 and p-siRNA-2 induced apoptosis in SW480 cells. The apoptotic rates were increased to 17% and 35.6% after treating with p-siRNA-1 and p-siRNA-2 for 48 hours, respectively, as compared to only 3% in the control group (Figure 3). Cleavage of caspase-3 is a hallmark of apoptotic cell death. Therefore, we determined the expression of caspase-3 in SW480 cells after treating with p-siRNA-1 and p-siRNA-2 for 48 hours by Western blot analysis. The data clearly showed the cleavage of caspase-3 (Figure 3C).
Figure 3: Induction of apoptosis in SW480 cells by periostin siRNAs. A, Silencing periostin expression induced apoptotic cell death in SW480 cells. The SW480 cells were transfected with control GFP lentiviral particles (a) p-siRNA-1 (b), p-siRNA-2 (c) for 48 hours, respectively, and then the cells were visualized under a fluorescent microscope. Green, lentiviral particle transfected cells; Red, annexin V stained cells (apoptotic cells). B, Data from Figure A are expressed as mean ± SEM, of three independent experiments, ap<0.01. C, SW480 cells were treated with p-control, p-siRNA-1 and p-siRNA-2 for 48 hours. Cell lysates were extracted and subjected to Western blot for the expression of caspase-3
Knockdown of periostin arrested the cell cycle at G0/G1 phase in SW480 cells
The cell cycle distribution of SW480 cells was determined by flow cytometry. As shown in Figure 4, the percentages of G0/G1 phase cells in the SW480 cells were significantly increased to 57.1 ± 3.0% and 55.8 ± 2.7% (vs 37.2 ± 3.9 in control group) after treatment with p-siRNA-1 and p-siRNA-2, respectively; with a corresponding decrease in G2/M phase cells from 26.9 ± 2.9% to 10.3 ± 1.9% and 11.1 ± 2.0, respectively.
Figure 4: Effects of cell cycle distributions in SW480 by periostin siRNAs. A, Flow cytometry analysis of cell cycle distribution in SW480 cells treated with p-siRNA-1 and p-siRNA-2, respectively, for 48 hours; B, Data were expressed as mean ± SEM. n=3, *p<0.01 compared with the control
Discussion
Colorectal cancer represents the 3rd leading malignant disease worldwide (Herszenyi et al., 2010; Hwang H, 2013). Progress in multidisciplinary treatment had greatly improved the outcomes of patients diagnosed with early stage of the disease, but the outcomes of those diagnosed with advanced stage of the disease remains still very poor (Dattatreya, 2013). Better prognosis of colorectal cancer patients depends on early detection of the disease. Primary screening methods in colorectal cancer include fecal occult blood test and tumor markers test, but none is verified sensitive and precise enough for the diagnosis of the disease. Therefore, there is an urgent need to identify novel screening- and prognosis-related biomarkers.
Periostin, a cell adhesion protein, is overexpressed in the tumor tissues of a variety of human tumors (Kudo et al., 2007). Several recent studies demonstrated that serum levels of periostin of lung cancer, hepatocellular carcinoma, cholangiocarcinoma, breast cancer, ovarian cancer, and colon cancer were up-regulated (Kudo et al., 2007; Ruan et al., 2009). Cumulative findings suggest that periostin might be a potential biomarker in cancer diagnosis. In the present study, we investigated both periostin expressions in colorectal cancer tissues and serum. We found that expression of periostin was up-regulated in colorectal cancer tissues and higher preoperative serum periostin levels in colorectal cancer were correlated to clinical stage and distant metastasis of the disease. Further in vitro studies indicated that SW480 human colon carcinoma cell expressed high level of periostin. We found here for the first time that knockdown of periostin by siRNA could arrest the cells at G0/G1 phase and induce apoptotic cell death in SW480 cells.
Serum biomarkers can be used for risk stratification, diagnosis, goal-directed therapy, and outcome prediction to many diseases including cancer. Ben et al (2009) reported that periostin serum levels were elevated in colorectal cancer, and suggested that periostin might be a potential colorectal cancer biomarker. In the present study, we demonstrated higher preoperative serum periostin levels in colorectal cancer than in healthy control, which correlates well with Ben and colleagues' results. However, contrary to Ben and colleagues' results, we and also other group (Xiao et al., 2013) detected robust periostin expression in SW480 cells.
Periostin is frequently up-regulated in human cancers and implicated in cancer cell proliferation, invasion, and epithelial-mesenchymal transition (Norris et al., 2008). Previous studies indicated that periostin inhibits apoptosis in several cell lines including SGC-7901 gastric cancer cells (Li et al., 2013), A549 non-small-cell lung cancer cells (Hong et al., 2010), U2OS osteosarcoma cells (Liu et al., 2010) and A2780 ovarian cancer cells (Zhu et al., 2011). We were curious to know whether periostin also correlated with cell apoptosis in SW480 cells. We found that knockdown of periostin expression with siRNA-periostin lentiviral particles induced apoptosis in SW480 cell. Our results suggest that periostin might be a promising target of therapeutical intervention for colorectal cancer. Further investigation is needed to explore the detailed molecular mechanism underlying periostin-induced apoptosis in SW480 cells.
Conclusion
Periostin was up-regulated in colorectal cancer tissues, and is closely related to stage of disease and distant metastasis. Knockdown of PERIOSTIN arrested the cell cycle at G0/G1 phase and induced apoptotic cell death in the SW480 cells. Therefore, periostin may be a promising prognostic and therapeutic target for colorectal cancer.
References
American Cancer Society. Cancer facts & figures for African Americans, 2013-2014.
Ben QW, Zhao Z, Ge SF, Zhou J, Yuan F, Yuan YZ. Circulating levels of periostin may help identify patients with more aggressive colorectal cancer. Int J Oncol. 2009; 34: 821-28.
Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet 2014; 26; 383: 1490-502.
Conway SJ, Molkentin JD. Periostin as a heterofunctional regulator of cardiac development and disease. Curr Genomics. 2008; 9: 548-55.
Conway SJ, Izuhara K, Kudo Y, Litvin J, Markwald R, Ouyang G, Arron JR, Holweg CT, Kudo A. The role of periostin in tissue remodeling across health and disease. Cell Mol Life Sci. 2014; 71: 1279-88.
Dattatreya S. Metastatic colorectal cancer-prolonging overall survival with targeted therapies. South Asian J Cancer. 2013; 2: 179-85.
Herszényi L, Tulassay Z. Epidemiology of gastrointestinal and liver tumors. Eur Rev Med Pharmacol Sci. 2010; 14: 249-58.
Hong L, Sun H, Lv X, Yang D, Zhang J, Shi Y. Expression of periostin in the serum of NSCLC and its function on proliferation and migration of human lung adenocarcinoma cell line (A549) in vitro. Mol Biol Rep. 2010; 37: 2285-93.
Horiuchi K, Amizuka N, Takeshita S, Takamatsu H, Katsuura M, Ozawa H, Toyama Y, Bonewald LF, Kudo A. Identification and characterization of a novel protein, periostin, with restricted expression to periosteum and periodontal ligament and increased expression by transforming growth factor beta. J Bone Miner Res. 1999; 14: 1239-49.
Hwang H. Colorectal cancer screening among Asian Americans. Asian Pac J Cancer Prev. 2013; 14: 4025-32.
Izuhara K, Arima K, Ohta S, Suzuki S, Inamitsu M, Yamamoto KI. Periostin in Allergic Inflammation. Allergol Int. 2014; 2014.
Khan M, Zheng B, Yi F, Rasul A, Gu Z, Li T, Gao H, Qazi JI, Yang H, Ma T. Pseudolaric Acid B induces caspase-dependent and caspase-independent apoptosis in u87 glioblastoma cells. 2012; Evid Based Complement Alternat Med. 2012.
Khan M, Qazi JI, Rasul A, Zheng Y, Ma T. Evodiamine induces apoptosis in pancreatic carcinoma PANC-1 cells via NF-κB inhibition. Bangladesh J Pharmacol. 2013; 8: 8-14.
Khoshbaten M, Pishahang P, Nouri M, Lashkari A, Alizadeh M, Rostami-Nejad M. Diagnostic value of fecal calprotectin as a screening biomarker for gastrointestinal malignancies. Asian Pac J Cancer Prev. 2014; 15: 1667-70.
Kudo Y, Siriwardena BS, Hatano H, Ogawa I, Takata T. Periostin: Novel diagnostic and therapeutic target for cancer. Histol Histopathol. 2007; 22: 1167-74.
Lech G, Slotwinski R, Krasnodebski IW. The role of tumor markers and biomarkers in colorectal cancer. Neoplasma 2014; 61: 1-8.
Li B, Wang L, Chi B. Upregulation of periostin prevents P53-mediated apoptosis in SGC-7901 gastric cancer cells. Mol Biol Rep. 2013; 40: 1677-83.
Liu C, Huang SJ, Qin ZL. Inhibition of periostin gene expression via RNA interference suppressed the proliferation, apoptosis and invasion in U2OS cells. Chin Med J (Engl). 2010; 123: 3677-83.
Luo JH, Zhou J, Gao Y. Correlation between periostin and SNCG and esophageal cancer invasion, infiltration and apoptosis. Asian Pac J Trop Med. 2013; 6: 516-19.
Norris RA, Moreno-Rodriguez RA, Sugi Y, Hoffman S, Amos J, Hart MM, Potts JD, Goodwin RL, Markwald RR. Periostin regulates atrioventricular valve maturation. Dev Biol. 2008; 316: 200-13.
Noura S, Yamamoto H, Ohnishi T, Masuda N, Matsumoto T, Takayama O, Fukunaga H, Miyake Y, Ikenaga M, Ikeda M, Sekimoto M, Matsuura N, Monden M. Comparative detection of lymph node micrometastases of stage II colorectal cancer by reverse transcriptase polymerase chain reaction and immunohistochemistry. J Clin Oncol. 2002; 20: 4232-41.
Price TJ, Segelov E, Burge M, Haller DG, Ackland SP, Tebbutt NC, Karapetis CS, Pavlakis N, Sobrero AF, Cunningham D, Shapiro JD. Current opinion on optimal treatment for colorectal cancer. Expert Rev Anticancer Ther. 2013; 13: 597-611.
Rose J, Augestad KM, Cooper GS. Colorectal cancer surveillance: What's new and what's next. World J Gastroenterol. 2014; 20: 1887-97.
Ruan K, Bao S, Ouyang G. The multifaceted role of periostin in tumorigenesis. Cell Mol Life Sci. 2009; 66: 2219-30.
Sorocos K, Kostoulias X, Cullen-McEwen L, Hart AH, Bertram JF, Caruana G Expression patterns and roles of periostin during kidney and ureter development. J Urol. 2011; 186: 1537-44.
Takeshita S, Kikuno R, Tezuka K, Amann E. Osteoblast-specific factor 2: cloning of a putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem J. 1993; 294: 271-78.
Wu G, Wang X, Zhang X. Clinical implications of periostin in the liver metastasis of colorectal cancer. Cancer Biother Radiopharm. 2013; 2013.
Xiao ZM, Wang XY, Wang AM. Periostin induces chemoresistance in colon cancer cells through activation of the PI3K/Akt/survivin pathway. Biotechnol Appl Biochem. 2013; 2013.
Zhu M, Saxton RE, Ramos L, Chang DD, Karlan BY, Gasson JC, Slamon DJ. Neutralizing monoclonal antibody to periostin inhibits ovarian tumor growth and metastasis. Mol Cancer Ther. 2011; 10: 1500-08.