Ethylmethylhydroxypyridine succinate, acetylcysteine and choline alphoscerate improve mitochondrial function under condition of cerebral ischemia in rat
Abstract
The study was devoted to see the effect of ethylmethylhydroxypyridine succinate, acetylcysteine and choline alphoscerate on the mitochondrial function under condition of experimental cerebral ischemia in rat. An integrated approach was used with the assessment of changes in the respirometric and antiapoptotic functions of mitochondria. It was found that the application of the objects under study contributed to a decrease in the intensity of glycolysis and the restoration of aerobic glucose metabolism with an increase in ATP synthesis, as well as a decrease in the dissociation of reactions in mitochondrial complexes I, II, IV and V. In addition, the introduction of the studied drugs reduced the intensity of caspase-dependent and caspase-independent apoptosis reactions, which, together with the restoration of mitochondrial respiratory function, contributed to the decrease in the size of the necrosis zone of the brain.
Introduction
Ischemic states of the brain is the second leading cause of death and primary disability of the population (Kim and Lee, 2018). In the pathogenesis of ischemic stroke, a significant role is given to the energy deficit associated with mitochondrial dysfunction (Kumar et al., 2016). It is known that the disconnection of the mitochondrial respiratory chain reactions leads to an intensification of oxidative stress, activation of glycolysis with a significant decrease in the ATP production (Di Meo et al., 2016). Further, a decrease in the cell's energy potential leads to the activation of the caspase-dependent pathway of apoptosis (effector caspase - caspase 3) (Yang et al., 2018). In addition, massive influx of Ca2+ ions into the cell, which develops as a consequence of glutamate excitotoxicity, contributes to the formation of mitochondrial pores of variable permeability, as a result of which the caspase-independent pathway of apoptosis is activated, coupled with cytochrome C and apoptosis-inducing factor release. As a result, the initiation of a cascade of apoptotic events leads to the accelerated death of neurons in the zone of ischemic (penumbra) and an increase in the necrotic area (Li et al., 2008). Thus, it can be assumed that a targeted effect on the mitochondria of the cell will eliminate the energy deficit and prevent the development of apoptosis reactions, which will ensure the preservation of neuronal integrity in conditions of cerebral ischemia.
Ethylmethylhydroxypyridine succinate is a water-soluble derivative of 3-oxopyridine, which has a wide spectrum of pharmacological activities including antioxidant, cerebroprotective and endotheliotropic activities (Balashov et al., 2007). Currently, the drug has been successfully used in the treatment of post-stroke conditions, coronary heart disease and heart failure (Sidorenko et al., 2011). Acetylcysteine also has significant therapeutic potential. The antioxidant, anti-inflammatory, antimicrobial, and anticarcinogenic properties of acetylcysteine are investigated (Pei et al., 2018). Choline alphoscerate is a structural phospholipid of cell membranes, promotes the accumulation of choline in the brain and has a cerebroprotective effect, contributes to restore cognitive functions in the post-ischemic period (Colucci et al., 2012). Thus, based on the types of pharmacological activity inherent in these drugs, it can be assumed that the use of ethylmethylhydroxypyridine succinate, acetylcysteine and choline alphoscerate is able to stabilize the function of mitochondria in condition of cerebral ischemia.
Materials and Methods
Biological model
The research is implemented on 50 male Wistar rats (220-240 g weight) obtained from the vivarium PMPhI - a branch of FSBEI HE VolgGMU. The animals were kept in controlled conditions: ambient temperature 22 ± 2°С, relative humidity 60 ± 5%. The rats were accommodated in macrolone T-3 cage. The granulated non-coniferous wood fraction was used as the underlay. The rats received food and water ad libitum. The underlay and accessories for water changed once every 3 days.
Study design
Ethylmethylhydroxypyridine succinate, acetylcysteine and choline alphoscerate were used as the test-objects. The studied drugs were administered per os in doses of ethylmethylhydroxypyridine succinate - 30 mg/kg (Voronkov and Pozdnyakov, 2018), acetylcysteine - 150 mg/kg (Patel et al., 2014) choline alfoscerate - 150 mg/kg (Tayebati et al., 2017) after 1 hour reproduction of ischemia and then daily for 3 days. After that time, the rats were decapitated and the biomaterial was collected for further investigation. The study design is presented in Figure 1.
Figure 1: Study design
Model of focal cerebral ischemia
Focal cerebral ischemia was modeled by irreversible right-sided thermocoagulation of the middle cerebral artery under chloral hydrate anesthesia (350 mg/kg). The area below and to the right of the eye was depilated, an incision was made and the soft tissues were moved apart, exposing the scion of the zygomatic bone, which was removed. After that, trepanation gap was drilled and a thermocoagulator burned through the middle cerebral artery under its intersection with the olfactory tract. Later, as far as possible, the topography of soft tissues was restored. The suture was treated with 5% iodine solution (Bederson et al., 1986). The biomaterial was taken on the 4th day after the reproduction of focal ischemia.
Biomaterial sample preparation
Rat brain was used as a biomaterial for respirometric analysis. The rats were decapitated under chloral hydrate anesthesia (350 mg/kg), organs were collected, after which the biomaterial was homogenized in a mechanical Potter homogenizer in a selection medium in the ratio 1:5 (1 mmol EDTA, 215 mmol mannitol, 75 mmol sucrose, 0.1% bovine serum albumin solution, 20 mmol HEPES, with a pH of 7.2). The cell population was obtained by differential centrifugation, for which the obtained biogenic homogenate was centrifuged in the mode of 1,400 xg → 3 min at 4°C, after which the supernatant was transferred to 2 mL tube. Next, the resulting supernatant was centrifuged at 13,000 xg → 10 min and the supernatant (culture contains native mitochondria) was removed for analysis (Sullivan et al., 2007). The ELISA study was performed in the supernatant of rat brain tissue obtained by centrifuging the HEPES brain homogenate in the 10,000 xg mode for 5 min according to the recommendations of the manufacturer of the reagent kit for ELISA analysis.
Determining the size of the necrosis zone
The size of the necrosis zone was determined by the triphenyltetrazolium method. The brain was removed, cut off the cerebellum, divided the hemispheres. Both hemispheres were weighed, then separately homogenized and placed in cups with 10 mL of 1% solution of triphenyltetrazolium chloride in phosphate buffer (pH 7.4). The sample bottles were placed in a water bath for 20 min at 37°C. Next, the brain tissue was precipitated by centrifugation at 1200 xg for 10 min. The supernatant was removed and 3 mL of cooled chloroform was added to the precipitate. Shake it for 2 min. The chloroform formazan extract was obtained for 15 min at 4°С, shaking the mixture every 5 min for 30 sec. Centrifuged and measured the optical density (492 nm) against pure chloroform. The calculation of the necrosis zone was expressed as percentage of the total mass of the hemispheres:
Where, x is the size of the zone of necrosis as percentage of the total mass of the brain; ε1 is the optical density of the sample with an intact hemisphere; ε2 is the optical density of the sample with a damaged hemisphere; M1 is the mass of the intact hemisphere; M2 is the mass of the damaged hemisphere
Respirometric analysis
Analysis of the state of the respiratory function of mitochondria was carried out by the method of respirometry using the AKPM1-01L laboratory respirometer system (Alfa Bassens, Russia). The mitochondrial respiratory function was assessed by the change in oxygen consumption in the medium against the introduction of mitochondrial respiratory uncouplers. The last in the work was: oligomycin 1 µg/mL; 4 - (trifluoromethoxy) phenyl) hydrazono) malononitrile (FCCP- 1 µM); rotenone - 1 µM; sodium azide - 20 mmol. The oxidation substrates were glucose - 15 mmol; pyruvic acid - 10 mmol; malate - 1 mmol; succinate - 10 mmol; ascorbate - 2 mmol; ADP - 1 mmol; N, N, N ', N'-tetramethyl-1,4-phenylenediamine (TMPD - 0.5 mmol). The overall assessment of mitochondrial function was determined by the level of oxygen consumption in the medium after sequential addition of oligomycin, FCCP and rotenone to the medium, and the ATP-generating ability was determined (by the difference in oxygen consumption after the addition of FCCP and oligomycin); the maximum level of respiration (according to the difference in oxygen consumption after the addition of FCCP and rotenone) and the respiratory capacity (according to the difference in oxygen consumption after the addition of FCCP and the basal level of oxygen consumption). The activity of glycolysis processes was evaluated when glucose was used as an oxidation substrate during the registration of oxygen consumption under the conditions of sequential addition of glucose, oligomycin and sodium azide to the medium. The intensity of glycolysis (according to the difference in oxygen consumption after adding glucose and the basal level of oxygen consumption), glycolytic capacity (according to the difference in oxygen consumption after adding oligomycin and glucose) and glycolytic reserve (according to the difference in oxygen consumption after adding glucose and sodium azide) were determined. Additionally, the activity of complexes I, II, IV, and V of the mitochondrial respiratory chain was evaluated. The activity of complex I was determined by the difference in oxygen consumption after adding the malate/pyruvate and rotenone mixture to the medium. The activity of complex II was evaluated by the difference in oxygen consumption after adding succinate and oligomycin to the medium. The activity of complex IV was determined by the difference in oxygen consumption after adding the mixture of rotenone/TMPD/ascorbate and sodium azide to the medium. The activity of complex V was evaluated by the difference in oxygen consumption after adding rotenone and ADP to the medium. During the analysis, the biosample volume was 275 μL, and 25 μL of injected analyzers. Oxygen consumption was determined in ppm (Sauerbeck et al., 2011).
ELISA study
In this study, the concentration of apoptosis-inducing factor, caspase-3, and ATP was determined by ELISA in rat brain supernatant. We used standard kit for ELISA analysis (Cloud Clone Corp., USA). The course of the analysis corresponded to the instructions attached to each kit.
Statistical analysis
Statistical processing of the obtained results was performed using the stat-analysis package STATISTICA 6.0. Data are presented as mean ± SEM. Comparison of medium groups was performed using the ANOVA method with the post-test of Newman-Keuls at p<0.05.
Results
Results of respiratory analysis
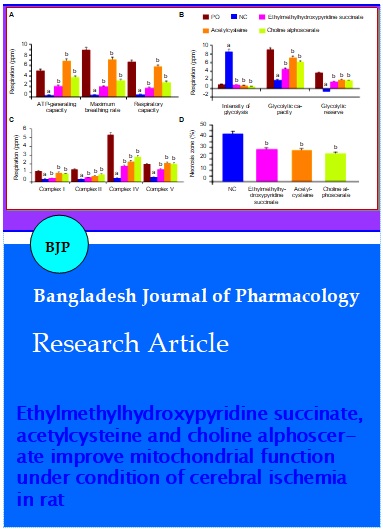
When conducting a respiratory analysis, it was found that in conditions of cerebral ischemia in the negative control rats relative to the pseudo-operated rats, there was a decrease in ATP-generating ability, the maximum level of respiration and respiratory capacity by 15.7 times (p<0.05); 20.6 times (p<0.05) and 16.1 times (p<0.05), respectively (Figure 2). At the same time, the negative control group compared to pseudo-operated rats showed an increase in glycolytic processes, as evidenced by an increase in the intensity of glycolysis, as well as a decrease in the glycolytic capacity and glycolytic reserve in rats deprived of pharmacological support relative to the pseudo-operated rats by 8.8 times (p<0.05); 4.6 times (p<0.05) and 5 times (p<0.05), respectively. The activity of the mitochondrial respiratory chain complexes I, II, IV, and V in the negative control group of rats was lower than the pseudo-operated rats by 3.5 times (p<0.05); 4.6 times (p<0.05); 12.5 times (p<0.05) and 3.7 times (p<0.05), respectively.
Figure 2: The effect of the test-objects on the change of mitochondrial respirometric function (A), process of glycolysis (B), mitochondrial respiratory chain complexes (C) and size of the brain necrosis zone (D) in rats under conditions of cerebral ischemia.
Note: astatistically significant relative to the pseudo-operated (PO) group of animals (p<0.05, Newman-Keuls criterion); bstatistically significant relative to the negative control (NC) group of animals (p<0.05, Newman-Keuls criterion)
The application of ethylmethylhydroxypyridine succinate in rats compared with the negative control rats showed an increase in ATP-generating ability, the maximum breathing rate and respiratory capacity by 6.6 times (p<0.05); 4.6 times (p<0.05) and 4.3 times (p<0.05), respectively (Figure 2). Also, in rats, when ethylmethylhydroxypyridine succinate was administered relative to the negative control rats, a decrease in glycolysis intensity was noted by 10.1 times (p<0.05) and an increase in glycolytic capacity by 2.3 times (p<0.05) glycolytic reserve in 2.2 times (p<0.05). The activity of complexes I, II, IV, and V of the mitochondrial respiratory chain in rats treated with ethylmethylhydroxypyridine succinate was higher compared with the negative control rats 1.2 times (p<0.05); 1.7 times (p<0.05) and 2.7 times (p<0.05), respectively.
When acetylcysteine was applied in rats, there was an increase (relative to the negative control rats) of ATP-generating ability, the maximum level of respiration and respiratory capacity by 21.7 times (p<0.05); 16.5 times (p<0.05) and 14.1 times (p<0.05), respectively. In addition, in rats with the introduction of acetylcysteine, the intensity of glycolysis was 12.4 times (p<0.05) lower, and the glycolytic capacity and glycolytic reserve respectively 3.7 times (p <0.05) and 2.7 times ( p<0.05) higher in relation to the negative control rats. At the same time, the activity of mitochondrial complexes I, II, IV and V increased in comparison with rats deprived of pharmacological support 2.9 times (p<0.05); 2.11 times (p<0.05); 5.4 times (p<0.05) and 4 times (p<0.05), respectively.
Against the background of the use of choline alfoscerate in rats relative to the negative control rats, an increase in ATP - generating ability was observed - 11.9 times (p<0.05), the maximum level of respiration - 7.3 times (p<0.05) and respiratory capacity - 7 times (p<0.05). Also in rats treated with choline alfoscerate relative to the negative control rats, there was an increase in glycolytic capacity and lycolytic reserve by 3.1 times (p<0.05) and 2.5 times (p<0.05), respectively, as well as a decrease in glycolysis intensity 19.5 times (p<0.05). The activity of the complexes of the mitochondrial respiratory chain in rats that were administered choline alfoscerate was higher relative to rats deprived of pharmacological support by 2.6 times (p<0.05) - complex I; 2.8 times (p<0.05) - complex II; 6.7 times (p<0.05) -complex IV and 3.9 times (p<0.05) - complex V.
ELISA analysis
The data obtained during the ELISA analysis are presented in Table I. The study found that the negative control rats relative to pseudo-operated rats showed an increase in the concentration of caspase-3 and apoptosis-inducing factor 10.6 times (p<0.05) and 4.9 times (p<0.05), respectively, while reducing the level of ATP by 3 times (p<0.05).
The use of ethylmethylhydroxypyridine succinate in rats compared to the negative control rats showed an increase in the concentration of ATP by a 2 times (p<0.05), and a decrease in the concentration of caspase-3 by 1.64 times (p<0.05) and apoptosis-inducing factor by 50% (p<0.05), respectively. When acetylcysteine was administered to rats, an increase in the ATP content was observed in relation to the negative control rats 2.2 times (p<0.05). The concentration of caspase-3 and apoptosis-inducing factor when using acetylcysteine decreased in comparison with the negative control rats 1.74 times (p<0.05) and 67% (p<0.05) respectively. The application of choline alfoscerate in rats showed an increase in the ATP content, as well as a decrease in the concentration of caspase-3 and apoptosis-inducing factor relative to the negative control rats by 2.5 times (p<0.05); 2 times (p<0.05) and 90.9% (p<0.05), respectively (Table I).
Size of the necrosis zone
In the course of this research unit, it was established that in the negative control rats the necrosis zone was 42.4 ± 2.4% (Figure 2). Against the background of the use of ethylmethylhydroxypyridine succinate, the necrosis zone decreased in comparison with the negative control rats by 48.4% (p<0.05). At the same time, with the application of acetylcysteine to experimental rats, this parameter decreased by 52.4% (p<0.05). With the use of choline alfoscerate, the area of the necrotized area of the brain was 69.7% (p<0.05) lower relative to the negative control rats.
Discussion
The study showed that the use of ethylmethylhydroxypyridine succinate, acetylcysteine and choline alphoscerate contributed to the preservation of mitochondrial function in conditions of cerebral ischemia in rats. The restoration of the functional activity of mitochondria was expressed in the normalization of ATP synthesis processes, by increasing the respiratory capacity and ATP-generating ability of mitochondria. Also on the background of the introduction of the studied objects to animals, limiting anaerobic glucose metabolism was observed with an increase in the energetically favorable aerobic oxidation pathway, which was confirmed by a decrease in glycolysis intensity, as well as an increase in glycolytic capacity and glycolytic reserve against the background of application of the studied objects.
Similar data were obtained in a study (Strifler et al., 2016) where the use of choline alfoscerate contributed to the restoration of mitochondrial membrane potential and reduced the manifestation of oxidative stress, including nitrite stress. For acetylcysteine, a positive effect on the change in mitochondrial function in ischemic stroke was also described, while it was found that the improvement of the mitohodrial potential was due to a decrease in the oxidative modification of cell structures that occurs when using acetylcysteine (Tardiolo et al., 2018). In addition, the introduction of acetylcysteine contributes to the suppression of the mitochondrial pathway of apoptosis, which may also indicate the restoration of mitochondrial function in the administration of acetylcysteine (Pei et al., 2018). The introduction of ethylmethylhydroxypyridine succinate also contributed to an increase in the intensity of cellular respiration in the mitochondrial isolate of the brain, while this effect is associated with the activation of succinate dehydrogenase (mitochondrial complex II) (Yasnetsov and Prosvirova, 2012).
Such changes in mitochondrial function under conditions of correction of experimental cerebral ischemia introduction ethylmethylhydroxypyridine succinate, acetylcysteine and choline alphoscerate may be associated with restoration of activity electron transport system of mitochondrial complex I, II, IV and V, which ultimately contributed to stabilization of oxidative phosphorylation process and, as a consequence, an increase in ATP synthesis (Ham and Raju, 2017). In addition, the restoration of aerobic metabolism has a positive effect on the course of apoptotic processes, both caspase-dependent and caspase-independent (Khosham et al., 2017). It is known that activation of the caspase-dependent pathway of apoptosis directly depends on the intracellular concentration of ATP, where with a decrease in the content of the latter, activation of proapoptotic Bid/Bax signaling molecules is observed, which initiate the caspase mechanism of apoptosis (Leist et al., 1997). In parallel with this process, due to the loss of mitochondrial membrane potential, an activation of the caspase-independent pathway of apoptosis is observed, with the result that enhanced apoptosis-inducing factor release causes activation of endonucleases followed by DNA sequencing (Natarajan and Becker, 2012). Thus, the cascade of apoptotic events is directly related to the function of mitochondria and the termination of the apoptotic signal may indicate the restoration of the function of mitochondria (Milasta et al., 2016). So in the course of this study, it was found that the use of ethylmethylhydroxypyridine succinate, acetylcysteine and choline alfoscerate contributed to a decrease in the concentration of caspase-3 and apoptosis-inducing factor, which in turn prevented the destruction of neurons in the ischemic penumbra zone and, ultimately, reduced necrosis size.
Conclusion
The study showed that ethylmethylhydroxypyridine succinate, acetylcysteine, and choline alphoscerate restore mitochondrial function under conditions of experimental cerebral ischemia. In addition, when using the studied objects, a suppression of the apoptotic signal was noted, which, together with the normalization of energy exchange, contributed to a decrease in the area of cerebral infarction.
Ethical Issue
The maintenance and all animal manipulations complied with the requirements of the European Convention for the Protection of Vertebrate Animals used for experiments and other scientific purposes (Strasbourg, 1986).
References
Balashov VP, Smirnov LD, Balykova LA, Gerasimova NG., Markelova IA, Kruglyakov PP, Talanova EV. Investigation of the role of the inducible isoform of NO synthase in the stress-protective activity of the ethylmethylhydroxypyridine succinate antioxidant. Russ J Cardiol. 2007; 2: 95-97.
Bederson JB, Pitts LH, Tsuji M, Nishimura MC, Davis RL, Bartkowski H. Rat middle cerebral artery occlusion: Evaluation of the model and development of a neurologic examination. Stroke 1986; 17: 472-76.
Colucci L, Bosco M, Rosario Ziello A, Rea R, Amenta F, Fasanaro AM. Effectiveness of nootropic drugs with cholinergic activity in treatment of cognitive deficit: A review. J Exp Pharmacol. 2012; 4: 163-72.
Di Meo S, Reed TT, Venditti P, Victor VM. Role of ROS and RNS sources in physiological and pathological conditions. Oxid Med Cell Longev. 2016; 2016.
Ham PB 3rd, Raju R. Mitochondrial function in hypoxic ischemic injury and influence of aging. Prog Neurobiol. 2017; 157: 92-116.
Khoshnam SE, Winlow W, Farbood Y, Moghaddam HF, Farzaneh M. Emerging roles of microRNAs in ischemic stroke: As possible therapeutic agents. J Stroke. 2017; 19: 166-87.
Kim JH, Lee Y. Dementia and death after stroke in older adults during a 10-year follow-up: Results from a competing risk model. J Nutr Health Aging. 2018; 22: 297-301.
Kumar R, Bukowski MJ, Wider JM, Reynolds CA, Calo L, Lepore B, Tousignant R, Jones M, Przyklenk K, Sanderson TH. Mitochondrial dynamics following global cerebral ischemia. Mol Cell Neurosci. 2016; 76: 68-75.
Leist M, Single B, Castoldi AF, Kühnle S, Nicotera P. Intra-cellular adenosine triphosphate (ATP) concentration: A switch in the decision between apoptosis and necrosis. J Exp Med. 1997; 185: 1481-86.
Li L, Peng L, Zuo Z. Isoflurane preconditioning increases B-cell lymphoma-2 expression and reduces cytochrome c release from the mitochondria in the ischemic penumbra of rat brain. Eur J Pharmacol. 2008; 586: 106-13.
Milasta S, Dillon CP, Sturm OE, Verbist KC, Brewer TL, Quarato G, Brown SA, Frase S, Janke LJ, Perry SS, Thomas PG, Green DR. Apoptosis-inducing-factor-dependent mitochondrial function is required for T cell but not B cell function. Immunity 2016; 44: 88-102.
Natarajan SK, Becker DF. Role of apoptosis-inducing factor, proline dehydrogenase, and NADPH oxidase in apoptosis and oxidative stress. Cell Health Cytoskelet. 2012; 2012: 11-27.
Patel SP, Sullivan PG, Pandya JD, Goldstein GA, VanRooyen JL, Yonutas HM, Eldahan KC, Morehouse J, Magnuson DS, Rabchevsky AG. N-acetylcysteine amide preserves mitochondrial bioenergetics and improves functional recovery following spinal trauma. Exp Neurol. 2014; 257: 95-105.
Pei Y, Liu H, Yang Y, Yang Y, Jiao Y, Tay FR, Chen J. Biological activities and potential oral applications of N-acetylcysteine: Progress and prospects. Oxid Med Cell Longev. 2018; 2018.
Sauerbeck A, Pandya J, Singh I, Bittman K, Readnower R, Bing G, Sullivan P. Analysis of regional brain mitochondrial bioenergetics and susceptibility to mitochondrial inhibition utilizing a microplate based system. J Neurosci Methods. 2011; 198: 36-43.
Sidorenko GI, Komissarova SM, Zolotukhina SF, Petrovskaya ME. The use of ethylmethylhydroxypyridine succinate in the treatment of patients with heart failure. Kardiologiia 2011; 51: 44-48.
Strifler G, Tuboly E, Görbe A, Boros M, Pécz D, Hartmann P. Targeting mitochondrial dysfunction with L-alpha glycerylphosphorylcholine. PLoS One. 2016; 11: e0166682.
Sullivan PG, Krishnamurthy S, Patel SP, Pandya JD, Rabchevsky AG. Temporal characterization of mitochondrial bio-energetics after spinal cord injury. J Neurotrauma. 2007; 24: 991-99.
Tardiolo G, Bramanti P, Mazzon E. Overview on the effects of N-acetylcysteine in neurodegenerative disease. Molecules 2018; 23: E3305.
Tayebati SK, Martinelli I, Moruzzi M, Amenta F, Tomassoni D. Choline and choline alphoscerate do not modulate inflammatory processes in the rat brain. Nutrients 2017; 9: 1084.
Voronkov AV, Pozdnyakov DI. Endothelotropic activity of 4-hydroxy-3,5-di-tret-butylcinnamic acid in the conditions of experimental cerebral ischemia. Res Result Pharm. 2018; 2: 1-10.
Yang JL, Mukda S, Chen SD. Diverse roles of mitochondria in ischemic stroke. Redox Biol. 2018; 16: 263-75.
Yasnetsov VV, Prosvirova EP. Investigation of the effect of Mexidol and cytoflavine on respiration of rat brain mitochondria. Bull New Med Tech. 2012; 19: 101-02.